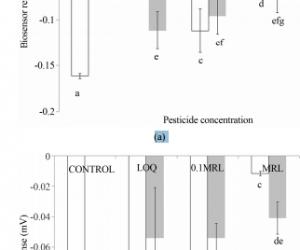
ABSTRACT Organophosphates belong to the most important pesticides used in agricultural practice worldwide. Although their analytical determinations are quite feasible with various conventional methods, there is a lack of efficient screening methods, which will facilitate the rapid, high-throughput detection of organophosphates in different food commodities. This study presents the construction of a rapid and sensitive cellular biosensor test based on the measurement of changes of the cell membrane potential of immobilized cells, according to the working principle of the Bioelectric Recognition Assay (BERA). Two different cell types were used, derived either by animal (neuroblastoma) or plant cells (tobacco protoplasts). The sensor was applied for the detection of a mixture of two organophosphate pesticides, diazinon and chlorpyrifos in two different substrates (tomato, orange). The pesticides in the samples inhibited the activity of cell membrane-bound acetylcholinesterase (AChE), thus causing a measurable membrane depolarization in the presence of achetylcholine (Ach). Based on the observed patterns of response, we demonstrate that the sensor can be used for the qualitative and, in some concentrations, quantitative detection of organophosphates in different substrates with satisfactory reproducibility and sensitivity, with a limit of detection at least equal to the official Limit of Detection (LOQ). The assay is rapid with a total duration of 3 min at a competitive cost. The sensitivity of the biosensor can be further increased either by incorporating more AChE-bearing cells per test reaction unit or by using cells engineered with more potent AChE isoforms. Standardization of cultured cell parameters, such as age of the cells and subculture history prior to cell immobilization, combined with use of planar electrodes, can further increase the reproducibility of the novel test. Keywords: Bioelectric Recognition Assay (BERA); Matrix Effects; N2a Cells; Organophosphates; Tobacco Protoplasts 1. Introduction Organophosphate insecticides have been used widely in agriculture and in household applications as pesticides due to their high insecticidal activity and relatively low persistence [1]. Their mechanism of action is the irreversible inhibition of acetylcholinesterase (AchE), a key enzyme in the recycling of the neurotransmitter acetylcholine (Ach) [2]. Organophosphates phosphorylate the serine hydroxyl group at the site of action of acetylcholine. They bind irreversibly, deactivating the esterase, resulting in accumulation of acetylcholine at the endplate. Decrease in plasma cholinesterase results in a decrease of cholinesterase activity in the central, parasympathetic, and sympathetic nervous systems. Accumulation of acetylcholine at the neuromuscular junction causes persistent depolarization of skeletal muscle, while neural transmission in the central nervous system is disrupted [3]. Long-term exposure to organophosphates has been associated with irritability, fatigue, headache, difficulties with memory and concentration and other neurophysiological abnormalities [4-6]. The conventional analysis of pesticide residues in food commodities is a labor intensive procedure, since it is necessary to cover a wide range of different chemicals, using a single procedure. Standard analysis methods include gas chromatography and high performance liquid chromatography to achieve the necessary selectivity and sensitivity for the different classes of compounds under detection [7]. As a consequence, current methods of analysis provide a limited sample analysis capacity, on a * day/instrument basis [8]. While the analytical determina- Corresponding author. Copyright © 2013 SciRes. CellBio 132 K. LOKKA ET AL. tion of the pesticide residues in an unknown sample will always be carried out using sophisticated methods based on conventional technology, rapid screening is the only solution for assuring food control by means of highthroughput detection of organophosphates in different food commodities. Providing novel solutions for food quality monitoring is also in accordance with the new EU and international regulations for minimal residue concentration in marketed food and agricultural products. Based on the inhibition of AchE and choline oxidase, many biosensors have been developed for the detection of organophosphorous and carbamate pesticides such as those described by Andres et al. [9], Choi et al. [10] and Andreou et al. [11]. It has been recently demonstrated that a successful pesticide assay could be based on a BERA platform, comprising electrically active cells (i.e. with an active transport of ions through the cell membrane) interfaced with microelectrodes which allow the capture of extracellular spikes or impedance changes associated with cellular response against the pesticide under detection [12]. More specifically, the inhibition of AchE by organophosphates and carbamate residues caused an increase of available Ach in the assay solution, which in turn caused the depolarization of the membranes of immobilized neuronal cells in a concentrationdependnet manner. In other words, the presence of the pesticides was detected by the degree of inhibition of cellular AChE, which is inversely associated with ACh concentration. Inhibition of AChE leads in increased excitatory ACh transmission and depolarization of the cell membrane, which was measured as a change of the sensor’s potential, due to changes in the concentration of electrolytes in the immediate vicinity of the working electrode. This novel type of biosensor was further included in a validation test against E.U. proficiency test samples [13]. The aim of the present study was to further develop this novel biosensor principle by incorporating, for the first time, plant (tobacco) protoplasts as organophosphate biorecognition elements. Their reliability as sensor components was compared to those of neuronal cells. In addition, both biosensor versions were applied for the detection of a mixture of organophosphate pesticides in different substrates, thus evaluating, also for the first time, possible matrix effects on the sensor’s performance. 2. Materials and Methods 2.1. Materials Diazinon (Diethoxy-[(2-isopropyl-6-methyl-4-pyramidinyl)oxy]-thioxophosphorane; CAS [333-41-5]; Mw = 304.35) and chlorpyrifos (O,O-diethyl O-3,5,6-trichloro- 2-pyridyl phosphorothioate; CAS [2921-88-2]; Mw = 350.59) (both purchased from Chem Device, West Chester, PA, USA) were used as standard organophosphate insecticides. Pesticide mixtures which contained 10 μM of each pesticide, were prepared daily in acetone solution. All other reagents were purchased from Fluka (Switzerland). 2.2. Cell Culture and Sensor Fabrication from Plant Protoplast and Neuroblastoma N2a Cells Plant protoplasts were isolated from tobacco (Nicotiana tabacum) leaves by preplasmolysing 0.5 g of them in 20 ml of CPW solution [14] supplemented with 0.7 M mannitol for one hour and then incubating them in 20 ml solution of the same composition and additionally supplemented with 3 mg pectinase (8.5 units/mg, from Aspergilus niger) and 2 mg cellulase (9.5 units/mg, from Trichoderma viridae) for 20 hours. One ml of protoplast solution contained 3.5 × 106 cells/ml. Mouse neuroblastoma (N2a) cell cultures were originally provided from LGC promochem (UK). Cells were cultured in Dulbecco’s medium with 10% heat-inactivated foetal calf serum (FCS), 10% antibiotics (streptomycin) and 10% l-glutamine. For incorporation into the biosensor, cells were detached from the culture vessel by adding trypsine/ EDTA for 10 min at 37˚C and further concentrated by centrifugation (6 min, 1200 rpm, 25˚C). For manufacturing consumable biosensors, 1 ml of plant or animal cells (at a density of 2.5 × 106 /ml) were mixed with 2 ml of 4% (w/v) sodium alginate solution and then the mixture was added drop wise, by means of a 22G syringe, in 0.8 M CaCl2. Each of the resulting calcium alginate beads had an approximate diameter of 2 mm and contained approximately 5 × 104 cells. The sensors were storable at room temperature in culture medium, under normal atmospheric conditions (i.e. non-CO2 enriched) for at least three weeks without any loss of performance. 2.3. Sample Preparation Organically-grown, pesticide-free tomato and orange fruits were used for the preparation of the assayed samples. Two groups of samples were prepared: The first group (fortified samples) comprised of blended fruits spiked with mixtures of diazinon and chlorpyrifos at various concentrations. Subsequently, 15 g of the blended fruit-pesticide mixture were homogenized in 30 ml acetone using an Ultra-Turrex homogeniser (model Silent Crusher, Heidolph) at 6000 rpm for one min. In following, 30 ml dichloromethane, then 30 ml petroleum ether were added and the mixture was stirred for one min. Then the mixture was centrifuged at 4000 rpm for 4 min at 20˚C. After removing 25 ml of the supernatant, the rest was left to dry in a waterbath, at 65˚C - 70˚C. Finally, the dry samples were redissolved in 5 ml methanol (10% v/v). The second group (matrix standards) was created Copyright © 2013 SciRes. CellBio K. LOKKA ET AL. 133 by spiking final dry samples, dissolved in methanol, with the pesticide mixtures. The concentration of each pesticide in the samples was calculated so that each pesticide was present either at the Limit of Quantification (LOQ), the Minimum Residue Level (MRL) or one-tenth thereof (MRL/10), according to the concentrations specified by the European Union Directive 839/2008 (Table 1). Control samples were created as the analytical sample but omitting spiking with pesticide mixtures. Dried control samples were also redissolved in methanol. 2.4. Assay Principle According to the working principle of the method, the presence of organophosphate compounds is detected by the degree of inhibition of cellular AChE, which is inversely associated with ACh concentration. ACh is an excitatory neurotransmitter (Hoang et al., 2007), the activity of which is regulated by AChE. Therefore, inhibition of AChE can lead in increased excitatory ACh transmission, which can be measured by the depolarization of the cell membrane. In other words, inhibition of AChE by pesticide residues in the sample will result to excessive stimulation of N2a cells by ACh, which will further lead to membrane depolarization above a predetermined threshold. Membrane depolarization events and associated electrolyte influx/efflux will reflect themselves on the sensor’s response as a change of the sensor’s potential, due to changes in the concentration of electrolytes in the immediate vicinity of the working electrode [12,13]. 2.5. Assay Procedure Each cell-bearing bead (cell sensor) was manually connected to a working electrode (the electrode was inserted through the entire length of the sensor without extruding from the opposite end) made from pure silver, electrochemically coated with an Ag/AgCl layer and having a diameter of 0.75 mm. The distance between working and reference electrode was 3 cm. Electrodes were connected to the recording device, which comprised the PMD-1608FS A/D card (Measurement Computing, Middleboro, MA) (Figures 1(a) and (b)). The software responsible for recording the signal and data processing was InstaCal (Measurement Computing). For each assay, the sensor system, comprising of the Table 1. LOQ and MRL points laid down by the European Union Directive 839/2008. Tomato Orange LOQ (ppm) MRL (ppm) LOQ (ppm) MRL (ppm) Diazinon 0.02 0.5 0.024 1 Chlorpyrifos 0.03 0.5 0.03 0.3 bead attached to the working electrode and a reference electrode, was immersed into each sample solution (200 μl). The sample solution comprised 150 μl of 50 mM Tris-aminomethane (Tris) buffer (pH 8), containing 0.5 mM acetylcholine iodide (Ach) and 50 μl of pesticide sample. The response of each sensor was estimated by recording the average change of the sensor potential for a period of 180 sec after sample application. 2.6. Data Analysis and Experimental Design Experiments were set up in a completely randomized design and each experiment was repeated three times. In each application, a set of six biosensors was tested against each individual sample. Correlations between the sensor’s response and pesticide concentrations were done using MS-Excel. Data means among different days were compared using Duncan’s multiple range test (with significance at p < 0.05). The effect of the extraction procedure and the concentration of the pesticides on the screening efficiency (SE%) in each sample was calculated individually for each sample, according to the following equation: average response to the fortified sample % 100 average response to the matrix standard ′ =SE 3. Results and Discussion 3.1. Response of the Sensor to Organophosphate Pesticides in Tomato Samples The response of the cell-based biosensor against different concentrations of a mixture of diazinon and chlorpyrifos in tomato samples is presented in Figure 2. The results Figure 1. (A) Schematic outline of the biosensor system. The graph shows the considerable increase of the response of the cellular biorecognition element to Ach after the addition of an organophosphate (chlorpyriphos) (red line). The insert (B) shows a Petri dish with consumable biorecognition elements (gel beads with immobilized cells). Copyright © 2013 SciRes. CellBio 134 K. LOKKA ET AL. of the assay using neuroblastoma as the sensor’s biorecognition elements are shown in Figure 2(a), while the results of the assay using plant protoplasts are shown in Figure 1(b). In the case of neuroblastoma N2a cells, when no pesticide was present in the sample (control (a) (b) Figure 2. Response of the cell-based biosensor against different concentrations of a mixture of diazinon and chlorpyrifos in tomato samples. The biosensor was based either on neuroblastoma cells (a) or tobacco protoplasts (b). Pesticide concentrations are expressed as the corresponding LOQ, MRL and MRL/10 values, according to Table 1. Sensor response is expressed as a change in the membrane potential of immobilized cells. (n = 6 replications (different sensors) for each sample and error bars represent standard errors of the average value of all replications with each sample). The white columns represent matrix standards and the grey columns fortified samples, as described analytically in the Materials section. Columns sharing a common letter are not statistically different (p ≥ 0.05). sample), a sensor response of −142 ± 2 mV was observed. The sensors responded to increasing pesticide concentrations by considerable positive increase of the sensor’s potential (Figure 2(a)). This observation is in accordance with the assay principle, where inhibition of AChE can lead in increased excitatory ACh transmission, which can be measured by the depolarization of the cell membrane (hence the shift of the sensor measurements to more positive values). The response of the sensor against matrix standards was quite reproducible (average variation = 12.7%) with the exception of the response against the highest concentration (MRL). Considering matrix standards only, a satisfactory correlation was observed between sensor response and total pesticide concentration (r2 = 0.9624, y = 0.042x – 0.1765). On the contrary, the response against the fortified samples was less reproducible, although a concentration-dependent pattern was again observed. The better reproducibility of the response of matrix standards vs. fortified samples could be due to the more homogenous distribution of the pesticides under detection in the matrix standards compared to the fortified samples. When plant protoplasts were used, the response to control solutions was less negative (−95 ± 1 mV) than with animal cells, indicating a lower state of membrane hyperpolarization of the immobilized cells. In this case, a correlation between pesticide concentration and biosensor response was observed only against matrix standards, not fortified samples (Figure 2(b)). In addition, a considerable variation of the sensor response was observed, much higher than for the N2a-based sensor. Depending on the concentration of the pesticides, the screening efficiency ranged from 42% to 141%. Thus, spiked pesticide concentrations lower than or equal to the LOQ were underestimated, while higher concentrations (0.1 MRL) were overestimated. 3.2. Response of the Sensor to Organophosphate Pesticides in Orange Samples The response of the cell-based biosensor against different concentrations of a mixture of diazinon and chlorpyrifos in orange samples is presented in Figure 3. The results of the assay using neuroblastoma as the sensor’s biorecognition elements are shown in Figure 3(a), while the results of the assay using plant protoplasts are shown in Figure 3(b). When no pesticide was present in the sample (control sample), a sensor response of −161 ± 3 mV (animal cells) or −119 ± 7 mV (plant protoplasts) was observed, i.e. steady-state membrane hyperpolarization was higher than for tomato controls. This was probably due to the different matrix effect: Mizayawa et al. [15] have previously reported that constituents of the essential oils of Citrus sp. considerably affected the activity of Copyright © 2013 SciRes. CellBio K. LOKKA ET AL. 135 (a) (b) Figure 3. Response of the cell-based biosensor against different concentrations of a mixture of diazinon and chlorpyrifos in orange samples. The biosensor was based either on neuroblastoma cells (a) or tobacco protoplasts (b). Pesticide concentrations are expressed as the corresponding LOQ, MRL and MRL/10 values, according to Table 1. Sensor response is expressed as a change in the membrane potential of immobilized cells. (n = 6 replications (different sensors) for each sample and error bars represent standard errors of the average value of all replications with each sample). The white columns represent matrix standards and the grey columns fortified samples, as described analytically in the Materials section. Columns sharing a common letter are not statistically different (p ≥ 0.05). membrane-bound AChE, thus affecting the potential difference along the membrane of the immobilized cells. Similarly to tomato samples, the response of the sensor dependent on the total pesticide concentration, although this effect was less pronounced using the plant cell-based version (Figure 3(b)). In addition, the response against orange samples was more reproducible than for tomato samples, especially against analytical orange samples. This lack of reproducibility (i.e. a considerable variation of response) was even more obvious when protoplastbased sensors were used to assay fortified samples, especially at the LOQ (Figure 3(b)). Depending on the concentration of the pesticides, the screening efficiency ranged from 38% to 350%, i.e. pesticide concentrations were either under- or overestimated, depending on the spiked concentration and the type of the biosensor, similarly to the assay of the tomato samples. On the other hand, and considering only matrix standards, a satisfactory correlation was observed between sensor response and total pesticide concentration for both animal cell-based (r2 = 0,8947, y = 0.0018x – 0,1454) and plant protoplast-based biosensors (r2 = 0.7923, y = 0.0321x – 0.1593). However, as indicated by the concentration-dependent screening efficiency for both types of biosensors, it was not possible to obtain a reliable quantitative response. By testing more pesticide concentrations in the future, it might be possible to identify a range of concentrations for each matrix X biosensor combination where the screening efficiency will be close to 100%, thus allowing for quantitative determination as well. Differences in the sensor responses between tomato and orange samples pesticides may be due to differences of the matrix effect. For example, tomato is rich in carotenoids, such as lycopene. It has been previously shown that carotenoids and their oxidation products promote gap junctional communication [16], therefore possibly affecting the electrophysiological behaviour of neuroblastoma cells. Matrix effects aside, differences in biosensor response to different batches of the same sample could be due to factors related to the cellular biorecognition element itself, such as the age of the cells (days elapsed between the detachment of cells from culture, subsequent immobilization and use of the biosensor) as well as subculture history prior to cell immobilization. It is strongly recommended to standardize these factors in order to minimize variability in biosensor response. As expected, biosensors based on neuroblastoma cells were more sensitive than protoplast-based ones, as demonstrated by the greater difference in response (cell membrane depolarization) compared to control samples. This is due to their membrane-bound AChE [17]. The occurrence of this enzyme on the membranes of tobacco protoplasts is also a fact, as first reported by Madhavan et al. [18] (though in guard, not mesophyll cells). However, the concentration of the enzyme on the plant cell membrane is considerably lower than on neuroblastoma cells, as corroborated by colorimetric assays by our group (unpublished data). Yet the sensitivity of the sensor sysCopyright © 2013 SciRes. CellBio 136 K. LOKKA ET AL. tem presented in this study is high enough to allow for the measurement of the pesticide-AChE interaction, even at the LOQ. The AChE-based higher level of response of neuroblastoma cells could be further exploited in order to increase the sensitivity of the novel assay, either by incorporating more AChE-bearing cells per test reaction unit, or by using cells engineered with more potent AChE isoforms. The first approach is currently under investigation in our laboratory with promising, though preliminary, results. The reproducibility of the system could be improved considerably by redesigning the cell-electrode interface. Quite recent experiments in our lab has shown that using screen-printed electrodes reduced variation in response against organophosphate pesticides in solution, though no matrix effects were yet assessed [19]. We plan to repeat the experiments described in the present study using planar electrodes. The novel assay has also been used for the detection of the avermectin abamectin and the pyrethroid α-cypermethrin [20], whereas various operational parameters, including assay temperature and electrode material were validated. A variation of the experimental approach described in the present study involved the seeding of N2a neuroblastoma cells on top of PEDOT electrodes treated with Nafion and Polylysine [21]. 4. Conclusion There are several areas of future work that would improve the utility of cell-based biosensors. Numerous applications would benefit from the development of parallel systems that allow for simultaneous measurements on multiple cell lines, thus improving both the breadth of sensitivity and the ability to discriminate or classify different groups of analytes. As demonstrated in the present study, the biosensor system can be used only for screening purposes, since it was not possible to achieve satisfactory quantitative determination. In addition, due to the vast differentiation of food commodities, a much wider range of samples, both in the context of matrix and residue composition should be tested before the novel system can be practically employed. Despite this, the novel biosensor for screening organophosphate pesticides offers a number of advantages over other conventional biosensor techniques, such as sensitivity and low cost, as well as the ability to monitor, in real-time, the presence of pesticide residues in food products. A particular advantage is the high speed of the assay (analysis time = 3 min), although this does not include the time required for sample extraction. However, we should mention that, for the practical application of the novel assay as a rapid screening tool, a single-or two-step extraction of the sample in an organic solvent would suffice. It must be emphasized that it is not meant to replace elaborate analytical methods but rather to assist the scaling up of food quality control, primarily designed to screen rapidly large amounts of agricultural products and food commodities for the presence of pesticide residues at the site of production (field), packaging, processing and/or sale. The application scope of the novel assay principle has already been extended to include complex organic contaminants [22] and mycotoxins [23]. Therefore, it represents a totally new generation of analytical instruments, enabling the implementation of food safety analysis by even minor users, such as small agricultural unions or food companies and can be potentially used by all parties involved in the chain of food production, processing and distribution, a market with a volume of 3.6 billion








论文发表 是一个专门从事期刊推广、论文发表、论文发表辅导的机构。融合收集数百家期刊杂志社征稿评职称,发表论文,以供广大作者免费阅览,以期待各位作者在短时间内掌握每种期刊征稿要求、审稿范围,在日常繁重的工作中短时间开心、放心快速投稿、在适合的时间拿到刊物,评上职称,不为挤不上评职称的末班车而烦恼。
本站主要整合如下评职称,发表论文:
教育论文发表
经济论文发表
科技论文发表
医学论文发表
计算机论文发表
文学论文发表
农业论文发表
学报论文发表
其它论文发表等,另期待更多的杂志编辑与本站联系文:
85782530(普刊发表)
82534308(核心发表)共同以正期刊界正刊之气,维护学术良好的氛围。
免责声明:本网所提供的信息资源如有侵权、违规,请及时告知!
论文网版权所有, 本站提供论文发表 论文投稿 发表论文 发表文章
文章只代表作者观点,并不意味着本站认同,部分作品系转载,版权归原作者或相应的机构;若某篇作品侵犯您的权利,请来信告知:lunww@126.com
中国论文网全权所属国际域名:www.lunww.com 中文域名:论文发表 技术支持:论文发表 国家双软认定单位 法律顾问:全民安律师事务所
期刊合作加盟咨询:5715378(加盟、投诉、建议) 论文发表咨询电话:18262951856 论文发表投稿邮箱:lunww@126.com
Powered by 论文发表 © 2010-2020

苏ICP备19023845号-4